|
綜述 | 川大華西醫院(IF:15.707):病毒組對結腸炎和結直腸癌的動態影響:免疫、炎癥、預防和治療(國人佳作)編譯:微科盟小木,編輯:微科盟茗溪、江舜堯。 微科盟原創微文,歡迎轉發轉載,轉載須注明來源《微生態》公眾號。 腸道微生物組包括一系列微生物基因組,如細菌組、病毒組、真菌組等。腸道微生物群在腸道免疫和疾病中起著至關重要的作用,例如炎癥性腸病(IBD)和結直腸癌(CRC);然而,其潛在的機制仍未被完全理解。闡明微生物群和炎癥之間的關系可能會極大地提高我們對病因、疾病進展、患者管理以及預防和治療發展的理解。在這篇綜述中,我們討論了腸道病毒(即共生病毒、致病性病毒和噬菌體)在結腸炎和結直腸癌的發生、發展和并發癥中的影響,以及它們在預防和治療方面的潛力。我們探討了腸道病毒和宿主免疫系統之間的相互作用對炎癥疾病和癌癥的嚴重程度的影響,包括腸道病毒與其他微生物和微生物產物之間的直接和間接相互作用。此外,通過具體臨床試驗的例子,解釋了病毒在腸道炎癥反應中的作用的潛在機制,以推斷潛在的治療靶點。鑒于迄今為止討論腸道病毒群的不同主題的文獻非常有限,我們相信這些廣泛的分析可能有助于深入了解IBD和CRC的分子發病機制,這可能可以幫助改進這些重要的人類疾病的治療方案。 論文ID 原名:Dynamic impact of virome on colitis and colorectal cancer: immunity, inflammation, prevention and treatment 譯名:病毒組對結腸炎和結直腸癌的動態影響:免疫、炎癥、預防和治療 期刊:Seminars in Cancer Biology IF:15.707 發表時間:2021.10.14 通訊作者:黃燦華&Min Wu 通訊作者單位:華西醫院生物治療國家重點實驗室、腫瘤中心,四川大學華西基礎醫學與法醫學院,成都生物治療協同創新中心;美國北達科他州立大學醫學與健康科學學院 DOI號:10.1016/j.semcancer.2021.10.004 綜述目錄 1 前言 2 病毒在人體腸道中的分布和豐度 3 與腸道炎癥和癌癥密切相關的病毒 4 可能與腸道炎癥和癌癥有關的病毒 5 以噬菌體為基礎的腸道炎癥和癌癥治療 6 治療CRC的溶瘤病毒 7 研究腸道病毒群的新興技術 8 挑戰與未來研究方向 附:縮寫 主要內容 根據國際癌癥研究機構(IARC)發布的GLOBOCAN 2020,結直腸癌(colorectal cancer, CRC)是2020年世界上第三大常見癌癥(發病率10%)和癌癥死亡的第二大原因(死亡率9.4%)。既往研究表明,慢性炎癥與結直腸癌的病因和發病機制有關,稱為結腸炎相關CRC(CAC)。炎癥性腸病(Inflammatory bowel disease, IBD),包括克羅恩病(Crohn’s disease, CD)和潰瘍性結腸炎(ulcerative colitis, UC),顯著增加了CAC的風險,例如,UC患者發生CAC的風險增加了2.4倍。CD患者的CAC發病率相似,但年齡低于健康人群。盡管CRC的分子機制尚不清楚,但一種可能性是與遺傳易感個體對腸道微生物群的免疫反應加重有關。反過來,微生物群也可能顯著影響宿主防御和炎癥反應,從而影響IBD和CRC的發展和進展。 腸道微生物群是一個復雜的生態系統,在人類健康和多種疾病的發病機制中發揮著重要作用。近幾十年來,隨著高通量、新一代測序技術和生物信息學的進步,數萬億細菌、真菌、病毒、寄生蟲和古生菌已被確認。已發現細菌(胃癌的幽門螺桿菌)和病毒(肝癌的乙肝和丙肝病毒以及宮頸癌的人乳頭瘤病毒)強烈影響癌癥發生(腫瘤發生)、進展和治療。結腸炎和癌癥微生物組研究幾乎完全集中在細菌成員基因組上。目前,在估計的1012種不同的微生物中,只有11種被認為是人類致癌物。越來越多的證據表明,僅靠某些微生物的存在和數量改變不足以致癌,但它們可以作為“第二擊”,促進致病性細菌感染、生態失調和宿主免疫調節的遺傳缺陷引起的炎癥和癌變。目前,大規模的臨床試驗正在測試微生物群對癌癥治療的影響,包括診斷、飲食調整和腫瘤內注射工程細菌。 幾十年的研究只發現了幾種直接導致癌癥的病毒,但似乎有很多病毒對宿主的免疫系統有復雜的影響。越來越多地認識到病毒感染參與IBD或CRC的情況。CRC是一個循序漸進的過程。病毒穩態失調可能引發或促進炎癥性疾病(如IBD),促進發育不良,最終導致癥狀嚴重和高死亡率的癌癥。已有研究表明,病毒直接參與炎癥和腫瘤的發生,因為它們有感染人類細胞和突變的能力。此外,病毒可以通過調節細菌群落的穩定性和組成間接起作用。因此,病毒被認為是潛在的模塊化生物療法。進一步了解病毒如何影響IBD和CRC,對于早期發現和預防癌癥早期階段之后的進展有很大的潛力,并顯著促進治療方法的發展。 在本文中,我們回顧了目前將病毒群與炎癥反應、結腸炎、CRC的發生和進展聯系起來的知識。本文總結了黏膜病毒的組成及其在健康和疾病中的直接作用(如結腸炎和CRC)。此外,本文還討論了病毒通過調節相關的細菌群落間接影響結腸炎和癌癥。最后,我們嘗試填補知識空白,并指出未來潛在的研究方向。 人類腸道病毒群包含腸道中自然存在的所有DNA和RNA病毒的基因組,包括噬菌體、真核病毒、古病毒和內源性逆轉錄病毒(圖1)。以往的研究主要集中在細菌(噬菌體)和真核病毒。除此之外,由于對腸道病毒群在人類中的作用了解有限,且病毒數據庫相對較小,所以對腸道病毒群的其他組成部分的研究還不夠深入。 噬菌體(phages)是細菌病毒中種類最豐富的一類,它通過免疫調節和殺菌作用來影響體內穩態。2003年首次公布了從單個健康個體中分離出來的人類腸道病毒群,每克至少有109-1010個病毒樣顆粒(virus-like particles, VLPs)(圖1)。腸道病毒群包括近1015個噬菌體,比細菌細胞多10倍,比人類細胞多100倍。以雙鏈DNA(dsDNA)噬菌體有尾病毒目(Caudovirales)和單鏈DNA(ssDNA)噬菌體微病毒科(Microviridae)為主的群居病毒具有高度的特異性、多樣性和穩定性。然而,這些噬菌體中的大多數仍未分類。核心噬菌體群落,包括crAss樣噬菌體,是人類腸道中最豐富和最廣泛的病毒(20-50%)。與非洲群體相比,西方社會crAss樣噬菌體的組成有顯著差異。此外,最近在孟加拉國和坦桑尼亞Laksam地區的個體中發現了一種獨特的流行噬菌體分類群(LAK噬菌體)。由于共生病毒參與宿主免疫培養和成熟,這些病毒的生態失調可能會導致IBD和癌癥,我們將在這篇綜述的后面進行討論。
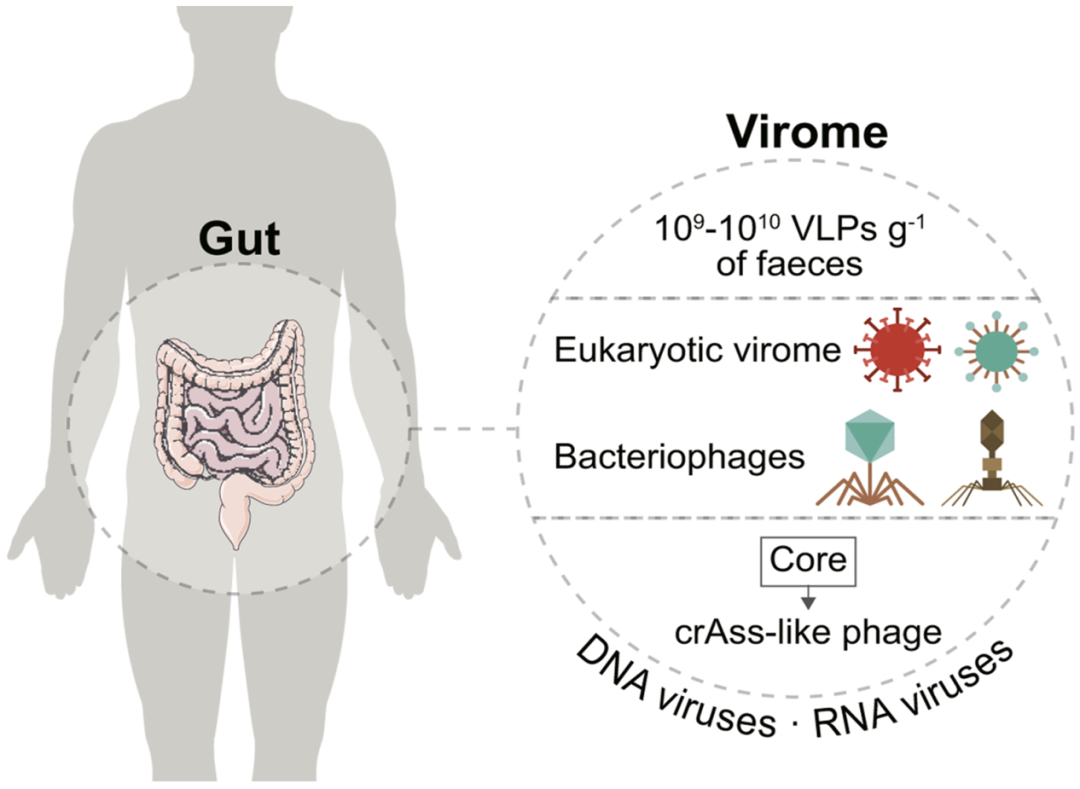
圖1 病毒在人體腸道中的分布。人類病毒群包含DNA和RNA兩種類型,人類每克毛重糞便中含有109-1010個病毒樣顆粒。病毒群的主要成分是真核生物、細菌(細菌噬菌體或噬菌體)、古菌病毒和內源性逆轉錄病毒。核心噬菌體群落是人類腸道中最常見的噬菌體,例如crAss樣噬菌體。該插圖是通過改編SMART (https://smart.servier.com)和Vecteezy(vecteezy.com)模板創建的。 多種腸道病毒可以感染人類,包括逆轉錄酶病毒、諾如病毒(norovirus)、輪狀病毒、腺病毒和皰疹病毒。例如,諾如病毒是全世界范圍內食源性腸胃炎的主要病因。小鼠諾如病毒(MNoV)在小鼠體內形成終身腸道感染,無癥狀個體可長期攜帶諾如病毒而無明顯疾病。腸道病毒感染可導致從無癥狀到中度、重度急性或重度慢性疾病的病理生理過程,可能會導致結腸炎和癌癥(圖2)。 在過去的20年里,隨著病毒宏基因組學的發展,病毒與腸道炎癥和癌癥之間的研究領域已經從檢測病毒粒子的存在發展到相互作用和病毒驅動的分子機制的研究(表1)。 圖2 宿主-病毒在腸道炎癥和癌癥中的相互作用。腸道病毒豐富度隨著IBD和CRC患者病情的加重而增加,而細菌多樣性和豐富度則降低,反映出疾病與細菌和病毒微生物的反向相關性。腸道炎癥的誘導或直接的遺傳毒性作用是微生物群誘導腸道癌變的主要機制。病毒可通過①直接感染腸道;②通過細菌間接影響宿主;③與宿主免疫系統相互作用并誘導免疫應答等途徑促進CRC的發生。該插圖是通過改編SMART(https://smart.servier.com)和Vecteezy(vecteezy.com)模板創建的。 表1 病毒與腸道炎癥和癌癥的關系。 IBD患者的噬菌體組成與健康對照明顯不同。與微病毒科(Microviridae)相比,CD或UC患者的有尾病毒目(Caudovirales)顯著增加,細菌豐富度和多樣性降低。在兒童CD患者的回腸活檢中也發現Caudovirales增多,但糞便中Caudovirales增多與UC發病無關。在另一項研究中,發現結腸中有皰疹病毒科(Herpesviridae)序列的患者顯示出人類內源性病毒基因表達增加和微生物多樣性增加。在早期診斷的未接受治療的IBD患者中,UC患者的Hepadnaviridae轉錄本數量高于CD患者和對照組,而Polydnaviridae和Tymoviridae含量較低。此外,與對照組相比,CD患者的Hepeviridae豐度增加,Virgaviridae豐度減少。在IBD患者中普氏棲糞桿菌(Faecalibacterium prausnitzii)通常減少,在IBD患者的樣本中發現F. prausnitzii噬菌體明顯高于家庭對照組,表明IBD中F. prausnitzii介導的死亡率增加。值得注意的是,特異性病毒感染可與IBD風險基因相互作用,改變IL-10或Atg16L1缺陷小鼠的腸道疾病,這表明某些種類的病毒可能會導致IBD的發生(圖3)。 圖3 腸道病毒對腸上皮細胞和宿主免疫細胞的潛在影響。在IL-10缺陷小鼠中,MNoV通過①微皺褶細胞穿過上皮屏障,②感染免疫細胞(如淋巴細胞、巨噬細胞和樹突狀細胞),并③引起干擾素反應(IFN-α,β)。在IL-10缺陷小鼠中,MNoV可④感染簇狀細胞,⑤誘導IFN-λ分泌,其對體內病毒的持續調控起關鍵作用。⑥持續性MNoV也可導致與IBD相關的Paneth細胞異常。⑦MNoV感染Atg16L1突變小鼠引發Paneth細胞異常,表現出腸道疾病。IBD, 炎癥性腸病; IEC, 腸上皮細胞; MNoV, 小鼠諾如病毒。該插圖是通過改編SMART(https://smart.servier.com)和Vecteezy(vecteezy.com)模板創建的。 CRC患者和健康對照組之間的病毒豐富度沒有顯著差異。然而,據報道,病毒失調與早期和晚期CRC有關。雖然病毒在CRC發病機制中的影響越來越大,但至今還沒有明確一致的共識結論。進一步嚴格的實驗和交叉隊列驗證是解決這一謎題的關鍵,建立或反證病毒與CRC的聯系,特別是在臨床患病率和病理生理學上。 在CRC標本中檢測到人乳頭瘤病毒(human papillomavirus, HPV)基因組,表明HPV的存在可能與患CRC的風險增加有關。然而,其他研究質疑HPV在結直腸癌發生中的作用,因為在結直腸癌標本中很少或沒有發現HPV DNA,這強調需要進一步的研究來闡明這種不一致。與HPV陰性組織相比,四種差異表達基因(WNT-5A, c-Myc, MMP-7和AXIN2)在HPV陽性CRC樣本中上調,這與一項早期的報告表明HPV與CRC發病機制相關一致。 有研究表明,人類巨細胞病毒(cytomegalovirus, CMV)優先感染CRC病變組織而非正常健康組織,這可能與CRC患者預后不良有關。這種現象可能與CRC細胞的增殖和進展有關,與對照組相比,CMV感染的CRC樣本中TLR2、TLR4、NF-κB、TNF-α表達升高,癌細胞系中Bcl-2、cox-2、Wnt/β-catenin表達升高。 多瘤病毒科(Polyomaviridae)的成員,主要是人類多瘤病毒2(稱為John Cunningham virus, JCV)被發現與CRC有關,表明JCV可能具有致癌作用。然而,在一些研究中,CRC樣本中很少或沒有檢測到JCV DNA。有人提出JCV通過以下方式促進結腸癌發生:首先,早期JCV蛋白T抗原(T-ag)被認為介導了病毒的致癌潛能,并與染色體不穩定性有關;其次,JCV可能負責誘導腫瘤抑制基因(如p53)的多態性和/或改變;最后,JCV可能會改變細胞行為(如遷移和侵襲),強調可能參與了PI3K/AKT、MAPK和/或Wnt/β-catenin通路。 在一項罕見的針對意大利人群的研究中,患有乙肝和丙肝病毒相關肝病的CRC患者表現出更好的5年無病生存期和較低的異時性肝轉移發生率,作者聲稱是“金屬蛋白酶抑制劑”假說,而不是病毒感染的直接影響。在另一項研究中,發現乙肝病毒(HBV)感染與結直腸癌風險增加密切相關,還有研究發現丙型肝炎病毒(HCV)感染與結直腸癌的發生沒有相關性。由于差異顯著,有必要進一步研究不同類型肝炎病毒、炎癥性疾病、CRC之間的相互關系。 相同的噬菌體在腸道炎癥環境中可能扮演著“雙刃劍”的角色。有尾病毒目噬菌體可顯著減少致癌性細菌的定植,提高易發生CRC的動物的存活率。同樣,CD患者中有尾病毒目噬菌體的豐度顯著增加。有必要分析這些復雜的相互作用,以測試病毒微生物是否可能在未來的研究中有治療益處。 因此,盡管有爭議,但這一領域的研究已經發現,一些真核病毒可以感染人類細胞,造成感染,觸發免疫反應,有時還會導致嚴重的疾病。未來的研究應探討病毒組成對特定腸道疾病(如IBD和CRC)的改變和影響。 腸道病毒首先暴露在細菌中,在宿主細胞(腸細胞或免疫細胞)中開始復制。動物感染模型已被普遍用于檢查腸道病毒和其他微生物之間的相互作用。目前,這些實驗已經通過使用抗生素處理和無菌小鼠或免疫缺陷小鼠和/或感染人類和小鼠病毒的年輕小鼠中進行。細菌微生物群對腸道病毒的有益作用直到最近才被認識到,這可能是因為許多研究涉及腹腔注射病毒而不是自然的口服途徑。 由于噬菌體和細菌在進化過程中進行著激烈的“軍備競賽”,噬菌體可能會改變細菌的微生物組,并通過復雜的機制在腸道生理和疾病中發揮作用,這需要進一步廣泛的闡明(圖4A)。首先,腸道噬菌體負責細菌群落之間的水平基因轉移(horizontal gene transfer, HGT),包括發病機制和抗生素耐藥性,這給控制細菌感染帶來了醫療負擔。其次,噬菌體的激活導致其細菌宿主的裂解和特定腸道細菌種類的豐度的變化。最后(但并非最不重要的),細菌裂解會釋放作為病原體相關分子模式(PAMP)和抗原的蛋白質、脂類和核酸,這可能觸發炎癥信號級聯,從而誘導細胞因子、細胞浸潤和組織損傷。最近的研究表明,病毒可以通過以下機制增強復制和傳播(圖4B):(1)細菌對病毒粒子的穩定作用。脊髓灰質炎病毒(Poliovirus, PV)通過糞口途徑傳播,可傳播至中樞神經系統。腸道細菌穩定病毒粒子并限制熱失活以增強PV復制和小鼠糞-口傳播。(2)細菌可增加宿主細胞附著。PV與脂多糖(LPS,革蘭氏陰性細菌表面的一種聚糖)和肽聚糖(革蘭氏陽性細菌細胞壁的主要成分)結合,增強病毒對宿主細胞的附著。(3)病毒結合LPS可誘導免疫耐受。小鼠乳腺腫瘤病毒(MMTV)是逆轉錄病毒科(Retroviridae)的一種,在人類中通過母乳從母親傳播給后代。MMTV與LPS結合,誘導宿主TLR信號和IL-10介導的免疫耐受,啟動病毒復制和傳播。(4)宿主IFN-λ可能受微生物調控。MNoV是杯狀病毒科(Caliciviridae)諾如病毒屬的一種,通過糞口途徑傳播,細菌可能通過調節IFN-λ反應促進MNoV的復制。 除了通過細菌間接影響宿主外,噬菌體還可以直接與宿主的免疫系統相互作用,引發免疫反應。越來越多的證據表明,在IBD或CRC患者腸道中,噬菌體與宿主細胞內免疫途徑相互作用并激活免疫應答。 共生病毒通過RIG-I-MAVS-IRF1-IL15軸和TLR3和/或TLR7 IFN-β通路抑制上皮內淋巴細胞,保護宿主動物免受右旋糖酐硫酸鈉(DSS)誘導的結腸炎(圖4C)。有尾病毒目噬菌體間接或直接刺激免疫反應并加重結腸炎。首先,噬菌體裂解細菌并釋放促炎產物;其次,噬菌體可以通過TLR9/MyD88依賴的方式促進淋巴結中CD8+和IFN-γ產生的CD4+ T細胞群的增加(圖4C)。被金黃色葡萄球菌(Staphylococcus aureus)或銅綠假單胞菌(Pseudomonas aeruginosa)噬菌體感染后,外周血單核細胞表現出轉錄反應,IL-1、IL-6、TNF的轉錄明顯增強。銅綠假單胞菌感染的人類傷口中的絲狀噬菌體觸發了免疫細胞中TLR3的激活和I型IFN的產生。反過來,這種類型的細胞因子(IFNα和IFNβ)抑制巨噬細胞產生TNF,從而損害吞噬作用和細菌清除,延遲傷口愈合。盡管在理解噬菌體與細菌的相互作用方面取得了這些進展,但要理解這種“軍備競賽”還為時過早。未來,更多的研究可能會集中在腸道噬菌體對影響噬菌體適應性和發病機制的細菌微生物群組成的影響上。 總的來說,病毒、細菌和宿主免疫系統之間的相互作用正在被更廣泛地研究。然而,在許多情況下,因果關系和分子機制仍然沒有被完全理解,應該從更廣泛的機制上進行探索。 圖4 跨界交互作用和機制。A,噬菌體通過宿主相關細菌直接或間接與宿主相互作用。這些相互作用可能影響宿主的遺傳變異,并對宿主健康產生明顯的影響。B,細菌增強腸道病毒復制和傳播的機制。C,病毒影響腸道的可能機制。①樹突狀細胞可通過RIG-I MAVS IRF1途徑感知病毒,刺激IL-15分泌,從而促進IELs增殖,抑制其凋亡。②駐留病毒被樹突狀細胞TLR3和TLR7識別,產生保護性IFN-β來抑制腸道炎癥。③噬菌體被內吞于樹突狀細胞中,激活B細胞和T細胞,刺激IFN-γ介導的TLR9依賴性免疫反應,從而加劇結腸炎。IEC, 腸上皮細胞; LPS, 脂多糖。該插圖是通過改編SMART(https://smart.servier.com)和Vecteezy(vecteezy.com)模板創建的。 以噬菌體為基礎的療法(也稱為噬菌體療法)已經存在了大約100年。60年前,傳染病的治療方法在大多數西方國家幾乎被拋棄,因為其結果難以預測,而且新發現的抗生素極大地改善了治療方法。然而,近幾十年來,抗生素的濫用導致了抗生素耐藥性的激增。由于缺乏有效的治療方法和細菌對抗生素耐藥性的迅速演變,噬菌體療法作為多藥耐藥細菌感染的替代方法重新受到人們的關注,主要是在非常嚴重的情況下。因此,越來越多的臨床報道和研究使用噬菌體治療致命性細菌感染或其他合并癥,如囊性纖維化(CF)和慢性阻塞性肺疾病(COPD)。 這并不奇怪,由于某些致病菌與IBD和CRC都有關聯,一些噬菌體相關療法的臨床試驗正在進行中,用于治療結腸炎和CRC。噬菌體治療可能在微生物操作方面具有優勢,因為噬菌體具有針對單一細菌的高度特異性。例如,噬菌體正在針對UC中的艱難梭狀芽胞桿菌(Clostridioides difficile)、CD中的粘附侵襲性大腸桿菌(Adherent-invasive E. coli, AIEC)和結直腸癌中的具核梭桿菌(Fusobacterium nucleatum)進行試驗。 粘附侵襲性大腸桿菌(AIEC)可能在CD發病機制中起一定的作用。Galtier等人從廢水中分離出三種針對AIEC的噬菌體,可以減少AIEC在回腸和結腸定植,減輕DSS誘導的小鼠結腸炎癥狀。這項工作為CD患者提供了一種新的治療選擇。一項2期雙盲、隨機、安慰劑對照臨床試驗(NCT03808103)招募了30例CD患者,以評估AIEC特異性噬菌體雞尾酒(EcoActive)對疾病活動、炎癥標志物和AIEC負荷的有效性。 具核梭桿菌(Fusobacterium nucleatum)參與CRC的發病機制。靶向F. nucleatum的噬菌體也進入了臨床試驗,利用這些病毒治療CRC并降低癌癥負擔。然而,這種單一的靶向也對利用噬菌體治療沒有特異性和有效噬菌體的新興菌株提出了挑戰,這些噬菌體可以通過從野生噬菌體中分離或通過工程設計生成多靶向通用噬菌體來開發。Gogokhia等人報道了從UC患者中分離的有尾病毒目噬菌體可以抑制致癌性粘附侵襲性大腸桿菌的生長,并抑制小鼠模型腸道腫瘤的生長。此外,噬菌體編碼一種解聚酶,使它們能夠降解生物膜并接近駐留的生物體。也可以設計具有額外治療優勢的噬菌體,例如,使用疊氮化物修飾的噬菌體連接伊立替康-葡聚糖納米顆粒治療CT26結直腸癌小鼠,可降低梭桿菌(Fusobacterium spp.)的水平,并有效抑制腫瘤生長。 以上研究強調,需要指出噬菌體療法在結腸炎和CRC中的潛在治療作用。噬菌體通常對細菌的靶標范圍很窄的主要限制可以通過設計多價、廣泛靶標的人工噬菌體來解決,或者儲存更多的野生噬菌體用于臨床應用。此外,擴大噬菌體療法在人類治療IBD和CRC中的應用還需要更大規模的隨機對照臨床試驗。 腫瘤病毒治療是以溶瘤病毒(Oncolytic viru, OVs)為基礎的免疫治療,通過調節腫瘤微環境(tumor microenvironments, TME)來逆轉免疫抑制狀態,進而刺激抗腫瘤免疫。OVs是一種天然的或經過基因改造的病毒,旨在靶向并殺死癌細胞,而不會對正常細胞造成明顯損害。目前,包括單純皰疹病毒(herpes simplex virus, HSV)、牛痘病毒(vaccinia virus, VAC)、腺病毒(adenovirus, AdV)、呼腸孤病毒(reovirus, RV)、麻疹病毒(measles virus, MeV)等在內的多種DNA和RNA病毒正迅速成為治療癌癥的新途徑。OVs目前通過基因修飾或與其他策略結合來優化,在不傷害健康細胞的情況下提供更大的特異性和對腫瘤的療效。表2總結了正在進行的CRC患者臨床試驗。 OVs可以作為癌癥疫苗,增加腫瘤特異性T細胞反應。OVs還可以裝備免疫刺激分子(如粒細胞-巨噬細胞集落刺激因子,GM-CSF),以提高其免疫激活特性。攜帶GM-CSF的OVs可促進樹突狀細胞(DC)的遷移和成熟,最終導致T細胞反應的啟動增強,如talimogene laherparepvec(T-VEC)和pexastimgene devacirepvec(Pexa-Vec或JX-594)。Talimogene laherparepvec (T-VEC)系統(HSV)是一種眾所周知的治療修飾病毒。T-VEC已經被美國食品藥品監督管理局(FDA)和歐洲藥品管理局批準用于治療轉移性黑色素瘤,T-VEC目前正在CRC臨床試驗中。Pexa-Vec是一種經過修飾的VAC,可編碼GM-CSF和β-半乳糖苷酶,以滅活病毒胸苷激酶基因,目前正在進行多項臨床試驗。 IL-12是Th1型癌癥免疫反應的主要調控因子,是另一種用于裝備OVs的細胞因子,如TBio-6517(I/II期)和ONCR-177(I期)。LOAd703 (Phase I/II)是一種雙臂ADV,由兩種腫瘤壞死因子受體(TNF)家族配體CD40L和4-1BBL組成,可刺激T細胞擴張、效應功能獲得、存活和T細胞記憶的發展。 Enadenotucirev (EnAd; ColoAd1)是一種復雜的嵌合病毒,由不同的腺病毒血清型重組而成,目前正在進行臨床研究。未修飾的病毒,如呼腸孤病毒Pelareorep (Reolysin),也正在用于CRC治療的臨床試驗。 此外,OVs可以與免疫檢查點抑制劑(immune checkpoint inhibitors, ICIs)或細胞毒性藥物結合,通過協同機制實現最有效的癌癥免疫治療,如Pexa-Vec與Tremelimumab(anti-CTLA-4)和Durvalumab(against PD-L1),TBio-6517與Pembrolizumab(anti-PD-1),ColoAd1與nivolumab(anti-PD-1),T-VEC與Atezolizumab(anti-PD-L1),OH2與HX008(anti-PD-L1),ONCR-177與Pembrolizumab。通過這些多管齊下的研究和臨床試驗,我們預計一些OVs可能能夠獲得FDA的批準,成功用于癌癥治療的臨床應用。 表2 使用溶瘤病毒治療CRC患者的臨床試驗。 注:5-FC, 5-氟胞嘧啶; 5-FU, 5-氟尿嘧啶; AdV, 腺病毒; CTLA-4,細胞毒性T淋巴細胞相關抗原4; dsDNA,雙鏈DNA; dsRNA,雙鏈RNA; GM-CSF,粒細胞-巨噬細胞集落刺激因子; HSV,單純皰疹病毒; i.a.,肝內動脈; i.t., 瘤內; i.v., 靜脈注射; MeV, 麻疹病毒; MSS-CRC,微衛星穩定型結直腸癌; PD-1,程序性細胞死亡蛋白1; PD-L1,程序性死亡配體-1; Ref, 參照; RV, 呼腸孤病毒; ssRNA, 單鏈RNA; TNBC, 三陰性乳腺癌; VAC, 牛痘病毒; VEGF, 血管內皮生長因子; T-VEC, talimogene laherparepvec。 1Clinicaltrials.gov標識符。2黑色素瘤、實體瘤、頭頸部鱗狀細胞癌、乳腺癌、晚期實體瘤、TNBC、CRC、非黑色素瘤皮膚癌、肝轉移瘤。來源: clinicaltrials.gov; 2021年5月評估。 高通量宏基因組測序技術的出現使我們能夠了解人類腸道噬菌體和各種病毒群體的復雜性和豐富性。然而,與腸道微生物組的細菌組成部分相比,腸道病毒群幾乎被忽視了,這主要是由于可用的病毒鑒定和分類工具有限;也可能是由于對細菌微生物組本身的重大興趣,所以病毒群的重要性在某種程度上被忽略了。此外,病毒以幾種不同的遺傳形式存在,它們的主要核酸(RNA或DNA)、鏈的特征(陽性或陰性)和鏈的數量(單鏈或雙鏈)不同。病毒的復雜性對庫的準備和測序策略提出了挑戰。目前估計只有1%的病毒群被測序,無法歸類的序列(分類或功能上)在60% - 90%之間(稱為病毒暗物質),有待鑒定。 VLPs組分可以使用透射電子顯微鏡(TEM)、宏基因組測序或高通量短讀長技術(Roche 454、Illumina平臺和Ion Torrent平臺)進行檢測。最近,兩種長讀長測序技術(Pacific Biosciences和Oxford Nanopore)被開發出來。這些技術可以幫助構建新的大型病毒基因組,獲得甲基化模式的信息,并在單個病毒粒子水平上研究種群結構。病毒宏基因組分析工作流程包括質量控制、reads的過濾和修剪、將reads組裝成contigs、去除細菌污染、在病毒數據庫中對病毒基因組進行比對以及下游分析。已經專門設計了一些軟件和數據庫來處理高通量病毒群測序數據(表3),這些工具應該在仔細考慮樣本類型和科學問題后使用。因此,一個關鍵的限制是是否有專業的生物信息學家和適當的硬件來執行如此龐大和耗時的分析。 除了病毒宏基因組測序外,未來的互補方法可能包括腸道病毒宏轉錄組學(RNA-seq)和病毒宏蛋白質組學。上述技術和其他未來發展可能會很好地塑造病毒群研究,以探索腸道病毒群的發病機制及其對人類的潛在有益影響,為設計有效的基于病毒群的腸道和結腸疾病治療方案提供見解。 表3 病毒宏基因組分析的選用方法和數據庫。 盡管取得了這些相對迅速的進展,但我們對腸道病毒特性的理解仍然存在重大差距。(1)雖然病毒參與結腸炎和CRC的發展已經越來越明顯,但目前的理論和觀察結果需要進一步的檢驗和驗證。(2)隨著許多新技術的出現,如核酸測序、組學分析和生物信息學管道,腸道病毒群的特性及其在病理條件發展中的作用仍處于早期階段。(3)病毒數據庫相對較小且不完整(與細菌基因組數據庫相比),限制了我們詳細分析健康和疾病中的粘膜病毒的能力。(4)有關人類健康的病毒學研究面臨著相當大的個體間差異,這些差異可能受到許多因素的影響,如年齡、性別、種族、地理、飲食以及樣本收集、儲存和處理。 總之,在病毒群研究、認識、疾病相關性和臨床應用/治療方面有許多重要進展,同樣也還有許多障礙。在噬菌體治療和OVs獲得廣泛臨床應用批準之前,必須適當解決上述障礙。盡管存在這些擔憂,但病毒療法可能值得探索,并且通過分子工程可能具有巨大潛力。 本文除了提供一個基本理念,并總結了病毒群與IBD和CRC的聯系外,還討論了噬菌體是一個很有前景的治療工具,以對抗人類炎癥性腸病和結直腸癌的致病菌。噬菌體治療的隨機、安慰劑對照試驗已經在IBD和CRC中進行。此外,迫切需要在病毒分離、宏基因組學、富集培養和生物信息學工具方面設計和發明新的方法,以提高我們未來定義和描述病毒的能力,需要進一步的大規模縱向和長期隨訪前瞻性研究,以及深入的實驗調查和驗證。該領域的關鍵方向是確定腸道微生物群與先天性免疫和適應性免疫之間的動態關系(如因果關系)。此外,噬菌體治療可與細菌或其他微生物組分(通過FMT、益生素和益生菌)一起應用,以改變毒性和免疫原性,有效地馴服人類加劇的炎癥疾病,改善人類健康。 附:縮寫 5-FC, 5-氟胞嘧啶; 5-FU, 5-氟尿嘧啶; AdV, 腺病毒; AIEC, 粘附侵襲性大腸桿菌; CAC, 結腸炎相關結直腸癌; CD, 克羅恩病; cDNA, 互補DNA; CMV, 巨細胞病毒; CRC, 結直腸癌; CTLA-4,細胞毒性T淋巴細胞相關抗原4; dsDNA,雙鏈DNA; dsRNA,雙鏈RNA; EnAd, Enadenotucirev;FAD,食品藥品監督管理局; gDNA, 基因組DNA; GM-CSF,粒細胞-巨噬細胞集落刺激因子; HBV, 乙型肝炎病毒; HCV, 丙型肝炎病毒; HPV, 人乳頭瘤病毒; HSV,單純皰疹病毒; i.a.,肝內動脈; i.t., 瘤內; i.v., 靜脈注射; IARC, 國際癌癥研究機構; IBD, 炎癥性腸病; ICI, 免疫檢查點抑制劑; IEC, 腸上皮細胞; JCV, John Cunningham病毒; LPS, 脂多糖; MeV, 麻疹病毒; MMTV, 小鼠乳腺腫瘤病毒; MNoV, 小鼠諾如病毒; MSS-CRC,微衛星穩定型結直腸癌; OV, 溶瘤病毒; PD-1, 程序性細胞死亡蛋白1; PD-L1, 程序性死亡配體-1; Pexa-Vec, pexastimogene devacirepvec; PV,脊髓灰質炎病毒; Ref, 參照; RV, 呼腸孤病毒; ssRNA, 單鏈RNA; VAC, 牛痘病毒; VEGF, 血管內皮生長因子; T-VEC, talimogene laherparepvec; TEM, 透射電子顯微鏡; TME, 腫瘤微環境; TNBC, 三陰性乳腺癌; UC, 潰瘍性結腸炎; VAC, 牛痘病毒; VEGF, 血管內皮生長因子; VLP, 病毒樣顆粒。 |